Symbiosis
Some reflections on the unit of relation
Published 29 October 2024 in Essays
30 mins read
Words by
Denise Araouzou
Drawings by
Olena Minyeyevtseva
Keywords
symbiosis, sympoiesis, emergence, systems thinking, lichen, language, symbiogenesis, social darwinism, modernity/coloniality
Why study symbiosis?
The biological process of different species living together over a prolonged period of time is called symbiosis. The word stems from Ancient Greek and is composed of σύν (sýn: together); and βίωσις (bíōsis: living). Unlike organisms performed extraordinary feats of cooperation throughout deep time so that humans may live with, and within such dazzling biodiversity today. The slimy, sticky, generative and at times confrontational process of getting get along is what I set out to understand through my study of symbiosis.
Symbiosis is found within and across micro-organisms (i.e cells, ecosystems, interspecies) as well as macro-organisms (i.e grassroots movements, political parties, social networks, orchestras, institutions). Although not always mutually beneficial, the evolutionary process of symbiosis demonstrates how vital interdependence is for life, not just to survive, but for more of it to continue to emerge. For all who are invested in building a future beyond the one dictated by necropolitical power (Mbembe, 2011), the words of scientists Margulis and Sagan (2001) serve as timeless reminder: “life did not take over the globe by combat, but by networking”.
In other words, to look at symbiosis with a curious mind may prove helpful in navigating the labyrinth of death encircling us. By focusing on what connects us, maybe the reasons we are divided become less important, less powerful. This is why lichen are the companion species for this essay.
Symbiosis

Mietoistenlahti, Finland. January 2024. Photo: Denise Araouzou
Lichens are composite organisms that defy tidy definitions and lines of descent. They comprise of diverse microbionts that belong to separate taxonomic kingdoms entangling algae or cyanobacteria, and fungi. The study of lichen revealed that across single-celled (microscopic) or multi-cellular (animals, plants, fungi) organisms are not as individual or autonomous as they may appear. In fact they are consortiums (Famintsyn, 1907), because two or more bacterial or microbial groups live symbiotically or, they can be explained as holobionts; that is when organisms are a host of many species (Margulis & Fester, 1991). Both rely on the communication and interaction among several species within and outside the organisms to evolve and survive.
The word symbiotismus was first coined by German plant physiologist Albert B. Frank (1839–1900) in 1877 during his study of lichen to describe “where two different species live on or in one another under a comprehensive concept which does not consider the role which the two individuals play but is based on the mere coexistence and for which Symbiosis is to be recommended” (translated in Sapp, 1994, p.6).

The algae or cyanobacteria will fuel lichen metabolism through photosynthesis, and the fungal partner, even following extreme cycles of wetting and drying, can re-organise its membranes and restart photosynthesis when moisture reappears. Artist, author and professor Laurie Palmer (2023) remarks in her book Lichen Museum that, “neither of the most prominent partners, the algae nor the fungus, would look or act, on their own, anything close to how they look and act together, nor would they thrive” (p. 35). This is precisely why lichen may be found across climates and biomes, from the frozen tundra to parched desert rocks (Tsing, 2012).
To be in symbiosis means to be involved in a generative and transformative process of becoming-with through a permanent, long-lasting intimacy of strangers (Margulis, 1998) that enables all entangled species and organisms to adapt to conditions that would have been otherwise unfavourable.
Symbiosis is not always a smooth encounter, nor is it a synonym for a mutually beneficial encounter. By the same token, it is also not always parasitic (extractive) and the conditions of the species’ interaction may change over time too. Nevertheless, the prolonged encounter of different species in the case of lichens, birds, pollinators, and more, allows both and all partners to thrive in environments where on their own, neither could survive.
Symbiogenesis
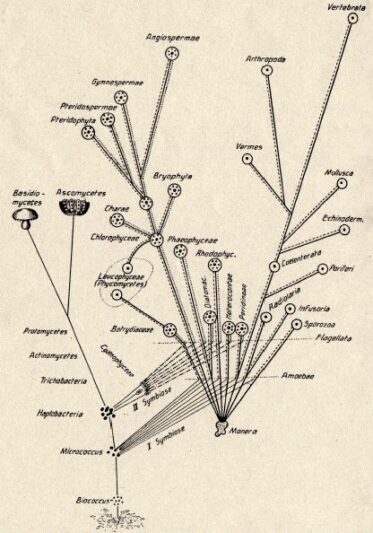
Konstantin Merezhkovsky, Symbiogenesis Tree of Life, 1910
Mitochondria are membrane-bound cell organelles (mitochondrion, singular) that generate most of the chemical energy needed to power the cell’s biochemical reactions. So the mitochondria’s purpose is to produce that energy. Some different cells have different amounts of mitochondria because they need more energy. So for example, the muscle has a lot of mitochondria, the liver does too, the kidney as well, and to a certain extent, the brain, which lives off of the energy those mitochondria produce. See here
Photosynthetic cells contain special pigments that absorb light energy. Different pigments respond to different wavelengths of visible light. Chlorophyll, the primary pigment used in photosynthesis, reflects green light and absorbs red and blue light most strongly. In plants, photosynthesis takes place in chloroplasts, which contain the chlorophyll. See here
Eukaryotes are organisms whose cells contain a nucleus and other membrane-bound organelles. See here
What happens when the prolonged encounter of two species not only allows for their survival, but for the emergence of new species?
In 1909, while Russian botanist Konstantin Merezhkovsky (1855-1921) was studying lichens on the Siberian tundra, he identified this process as symbiogenesis. Studies in symbiogenesis were occurring simultaneously as plant and marine biologists looking at micro and macro organisms became increasingly critical of the dominant Darwinian idea of evolution predicated on struggle and competition.
Even though symbiogenesis emerged as a key driver of evolution in the work of several scientists working across disciplines and geographies in the beginning of the early 20th century, Western mainstream zoocentric science did not recognise it as such until almost a century later (Hird, 2009). Evolutionary biologist Lynn Margulis wrote her first paper on symbiogenesis titled on The Origin of Mitosing [Eukaryotic] Cells in 1966; it was rejected by fifteen scientific journals before it was finally published in 1967. She became symbiosis’s most ardent promoter, attributing the bacterial origins of human mitochondria and photosynthetic chloroplasts to the symbiogenesis of eukaryotic cells and disproving, with genetic evidence, that organisms are closed, individual, self-sufficient systems.
Since the origin of nucleated cells – the latest major evolutionary step —evolution has only produced different variations of the same essential type of eukaryotic organisms. All fauna, flora, funga and many unicellular organisms are eukaryotes. One wild, but vital leap of the imagination is to imagine for a moment that trees, human mammals and mushrooms are variations of one another.
The origin of the four eukaryotic kingdoms according the Lynn Margulis’ Serial Endosymbiogenetic Theory. The evolutionary transition from prokaryotes to eukaryotes happened because of symbiogenesis.
Courtesy of Ricardo Guerrero.
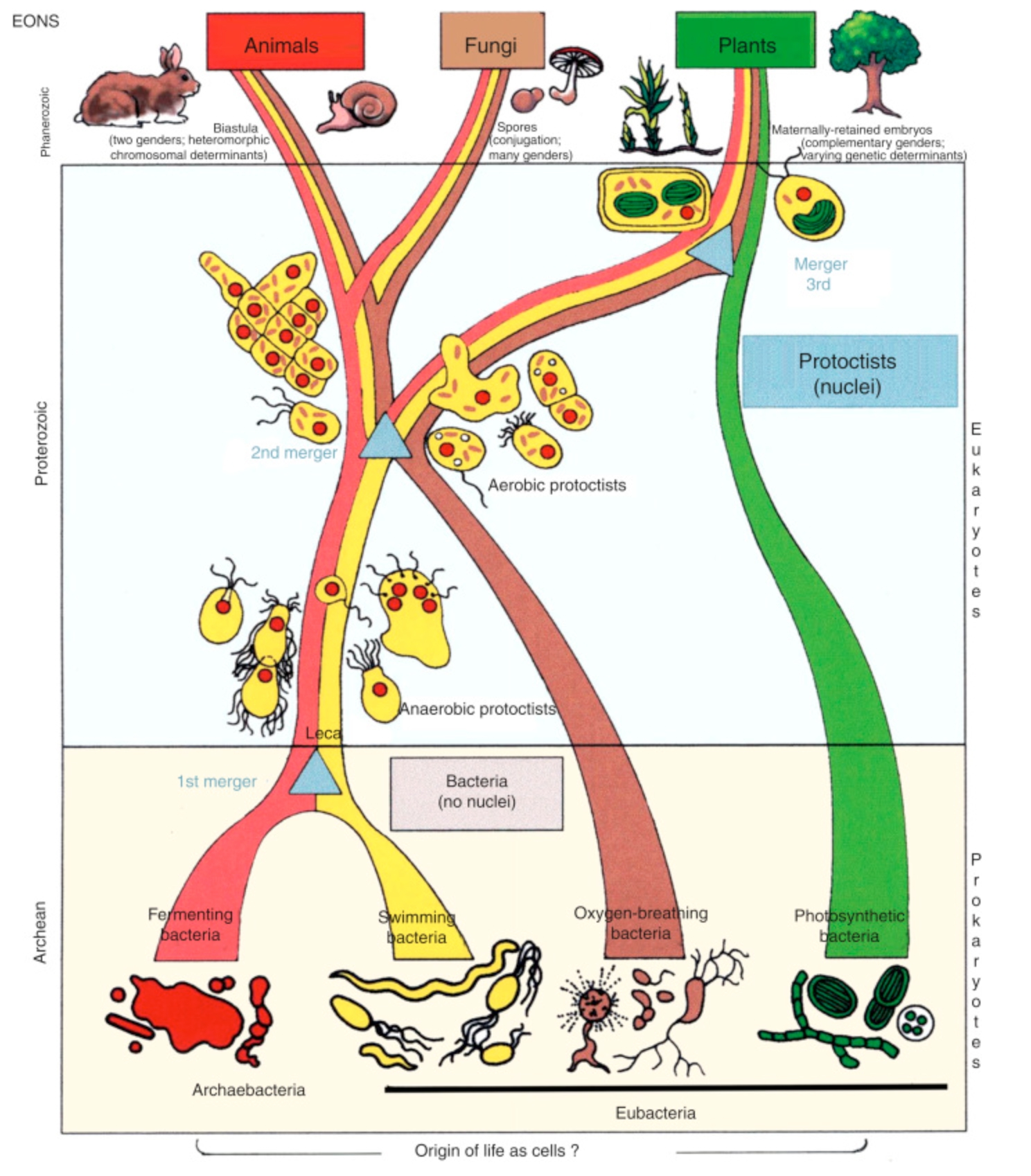
All organisms large enough for us to see are composed of once-independent microbes, teamed up to become larger wholes. As they merged, many lost what we in retrospect recognize as their former individuality. My theory of the symbiogenetic origin of plant, animal, and other cells with nuclei employs four provable postulates. All four involve symbiogenesis, incorporation, and body fusion by symbiosis.
Lynn Margulis (1998)

What stories we tell to tell other stories

Latinx professor and decolonial educator Machado de Oliveira Andreotti (2021) proposes the term modernity/colonialityto articulate planetary devastation in terms of layers. Rather than perceiving it as an inanimate object, or limiting it with a label that abdicates control over to whatever situations, configurations of power initiate these cycles (-cene stems from καινός, meaning ‘new’ in Greek), she extends her analysis of what she perceives to be a ‘multifaceted, living entity’ comprised of modernity and the processes of coloniality that it relies on to survive (incl. expropriation, extraction, exploitation, militarization, dispossession, destitution, genocides, and ecocides). She suggests it is “a worlding story, a complex adaptive living system that actively does things, including conditioning the habits of knowing and being of those whose lives and livelihoods are intertwined with it” (p.86).
The ongoing and increasingly alarming planetary shifts in climate, earth system functions and biodiversity extinction rates seem to continually slip through language. A plethora of definitions have been proposed within the Anglocentric academic world by various scientists, theoreticians and thinkers to make sense of this polycrisis – the Anthropocene proposed by Crutzen (2006) being the most popular and disputed term of all. Others include capitalocene (Moore, 2017); anthrobscene (Parikka, 2015); plasticene (Haram et al., 2020) and, plantationcene (Haraway, 2015). These neologisms reveal a few things. First, our incapacity to fully grasp undoings of planetary proportions; secondly, our urge to use words to feel out the limits and margins of our knowledge and purpose and thirdly, and most importantly, the limits of language to instigate radical action.
Margulis was not fond of applying biological processes to interpret social phenomena even though she did recognise their impact. However, in this essay I wish to explore how fundamental evolutionary theories and sciences have been shaping the material-discursive phenomena of modernity/coloniality. Even though Margulis was not fond of it, they are, politically, socially and economically deeply intertwined. Therefore, anyone aspiring to contribute to urgent eco-social transformation could begin by revisiting the stories we tell about how life came to be and recognise how these stories brought to the fore what we have to reckon with today. Amitav Ghosh guides us through the essence of this recognition in The Great Derangement: Climate Change and the Unthinkable (2016):
Recognition is famously a passage from ignorance to knowledge. To recognize, then, is not the same as an initial introduction. Nor does recognition require an exchange of words: more often than not we recognize mutely. And to recognize is by no means to understand that which meets the eye; comprehension need play no part in a moment of recognition. The most important element of the word recognition thus lies in its first syllable, which harks back to something prior, an already existing awareness that makes possible the passage from ignorance to knowledge: a moment of recognition occurs when a prior awareness flashes before us, effecting an instant change in our understanding of that which is beheld. Yet this flash cannot appear spontaneously; it cannot disclose itself except in the presence of its lost other. The knowledge that results from recognition, then, is not of the same kind as the discovery of something new: it arises rather from a renewed reckoning with a potentiality that lies within oneself (p.4).

The stories we tell about how life came to be should begin from recognising that we emerged from the living together of many. Symbiosis will now ‘spill out’ of the natural sciences and into philosophical and social science (re) considerations which shaped our understanding of agency, subjectivity and survival.
The seminal publication by Charles Darwin On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle of Life was published in 1859. Since then it has been appropriated to serve extractive capitalist, racist and oppressive means and ends. In the 20th century, between the 1930s and 1950s the so-called Modern Synthesis or New Synthesis (Huxley, 1942) combined Darwin’s theory of evolution with Gregor Mendel’s work on genetics into a joint mathematical framework. As a consequence, a great deal of complex systems, organisations and relations, including code fragments, genes, cells, organisms, populations, species, and ecosystems, were described as and reduced to mathematical equations of competition and struggle. Among many famous works inspired by this logic the work of British evolutionary biologist and zoologist Richard Dawkins’s The Selfish Gene (1976) stands out for the lasting impact his sociobiological analysis had on our contemporary social imaginary. He argues that all animals, plants, bacteria, and viruses are “survival machines, robot vehicles blindly programmed to preserve the selfish molecules known as genes” (p.xxi). In addition, the expression survival of the fittest he refers to in order to justify our inherently selfish behaviour is attributed to philosopher and sociologist Herbert Spencer who, inspired by Darwin, wrote Principles of Biology(1864) in which he outlines correlations between Darwin’s theory of natural selection and his laissez-faire economic theory.
It matters what matters we use to think other matters with; it matters what stories we tell to tell other stories with; it matters what knots knot knots, what thoughts think thoughts, what descriptions describe descriptions, what ties tie ties. It matters what stories make worlds, what worlds make stories.
Haraway (2016)

Neo-Darwinists not only argued that the selfish gene and survival of the fittest is a law of nature but also attributed the creation of new species to chance variations/mutations during gene reproduction, while disregarding the evolutionary mechanism of symbiogenesis and reducing fitness to the number of offspring an organism has. This misreading of Darwin’s theory of evolution lay the foundations for the theory of Social Darwinism which taught us that greed and competition are inherent in all evolutionary processes, hence, life. Since then, this narrative fanned the flames of the capitalistic zeitgeist and the justification of racism, interspecies oppression, extractivism, colonialism, eugenics and genocide – all in the name of becoming the ‘fittest’. Even though these crimes were intrinsic to imperialist expansionism well-before the 19th century, once the they were reinforced by modern science they became entrenched in Western society, across education, geopolitics, technology and more (Hird, 2009; Haraway, 2016).
Language matters, after all
Margulis (1995) reminds us that “Symbiosis has nothing to do with cost or benefit. The benefit/cost people have perverted the science with invidious economic analogies […] the language of life is not ordinary arithmetic and algebra; the language of life is chemistry” (p.135). She also explains to all those who may resort to the powerful allegory of symbiosis, like myself, that “Symbiosis is not a social relationship because social relationships refer to associations between members of the same species. To be symbiotic a relationship must refer to members of different taxa” (cited in Feldman, 2017). Yet, several scholars in the arts and sciences have since began to recognise that processes characteristic of symbiotic relations are not limited to the same species.
For example, scholar of Social Theory, Science and Technology Studies Maurizio Meloni admits in ‘How Biology Became Social, and What it Means for Social Theory’ (2014) that, “the traditional separation between the biological and the social has become increasingly difficult to define. […] sociology is becoming more open to biological suggestions, just at a time when biology is becoming more social” (p. 594). He attributes the ‘social rethinking of biology’ to three conceptual novelties:
Technical changes in evolutionary literature that have made possible a rethinking of the possibility of altruism and cooperation in evolution (prosocial view of evolution); Changes in neuroscience from the brain as an isolated data processor in neuroscience to a multiply connected device profoundly shaped by social influences (the social brain); Finally, and probably more importantly, changes in molecular biology, from the view of the gene as an autonomous master of development to the ‘reactive genome’ (cited in Keller, 2011; Griffiths and Stotz, 2013) of contemporary postgenomics (the socialized gene), a notion perfectly embodied in the new emerging field of molecular epigenetics (p. 594)
As Meloni (2014) encourages us to see how the boundaries between the social and the biological have become porous, Literature scholars Karpouzou & Zampaki (2023) prompt us to reflect how allegory may shape different imaginaries, they suggest that, “Symbiotic allegories and metaphors could pave a path to understanding and representing different real, potential, or fantastic agencies in literature and art” (p. 20). If the boundaries between the social, the biological and language dissolve, what if, they could also have ‘real, potential, or fantastic agencies’ in the ethics, philosophies, ontologies, epistemologies and values that (re)produce our social imaginaries? What if, symbiotic allegories could be the lens with which the complexity of relations in an entangled world is communicated, shared, spoken about? What if these ‘fantastic agencies’ help us recognise our very real interdependencies?
Among Margulis’ many accomplishments and contributions to evolutionary theory and science, perhaps the most important one is that she taught the world to recognise itself as becoming through/in relation. Gradually, her theories became recognised by mainstream science and to this day they still inform evolutionary studies. Even Dawkins finally admitted the importance of her contribution to science:
I greatly admire Lynn Margulis’s sheer courage and stamina in sticking by the endosymbiosis theory, and carrying it through from being an unorthodoxy to an orthodoxy. I’m referring to the theory that the eukaryotic cell is a symbiotic union of primitive prokaryotic cells. This is one of the great achievements of twentieth-century evolutionary biology. (Brockman, 1996)
LICHEN AS TEACHERS
The close and careful observation of patterns, characteristics and systems in ecosystems and more-than-human life have offered endless inspiration for several domains of human knowledge creation ranging across: agroecology, cybernetics, engineering and art to name a few. Laurie Palmer (2023) has spent a great deal of time studying and writing about lichen. She suggests these five qualities we can learn from lichens.
Their collectivity: a lichen is a symbiotic relationship between a fungus and some algae as well as a miniature ecosystem of many organisms cutting across taxonomic lines messing with the idea of individual identities of linear evolutionary trees and pointing towards the survival strategies of mutual aid.
Their site specificity: they constitute themselves an intimate relationship with their environment and can’t be separated from it reminding us that the environment is never a separate entity but we are part of it and it is in us.
Their slow pace: their long-term perspective yearly growth is generally measured in millimeters and some can live thousands of years if they’re understood to die at all. This is a very different time frame than the speed up of global capital.
Their refusal to be commodified: they can’t be grown in a lab and are too small and too slow to provide a steady source of anything for capital to feed on.
Their diversity: wild diversity of form, porosity and sensitivity to their environment makes them highly adaptable. (p. 13)
To apply these qualities in practice may sound like an metaphor as experiment, but they are actively implemented across several grassroots movements, activist circles and all kinds of collectives. But what about larger or more complex configurations? Does the notion of symbiosis translate into larger systems? For example, relations between the collective and the public, or a populated village and the surrounding landscape or a neighbourhood and the local fauna, flora and funga?
SYMPOIETIC SYSTEMS

M. Beth Dempster was an Environmental Studies Master student in 1998 when she coined the word sympoietic in her thesis ‘A self-organising systems perspective on planning for sustainability’. This word also stems from Ancient Greek and is composed of the prefix σύν (sýn: together) and ποιήσης (poíēsis: to make, poetry). She used it to define collectively producing systems that do not have self-defined spatial or temporal boundaries, where information and control are distributed among components, and the systems are evolutionary and have the potential for surprising change (p.30).
A system is a set of relations that describes some sense of connectedness. Even though, components and boundaries also contribute to the understanding of the kinds of systems, Dempster (1998) argues that it is the relations (my emphasis) among these components, and how they are established, that are even more critical for establishing a system, its behaviour, and its degree of complexity.
Prominent scholar in the field of science and technology Donna J. Haraway (2016) builds on the work of Dempster and explains that sympoiesis is “a carrier bag for ongoingness, a yoke for becomingwith, for staying with the trouble of inheriting the damages and achievements of colonial and postcolonial naturalcultural histories in telling the tale of still possible recuperation” (p. 125). They are the evolutionary systems from which the recuperation and survivability of living systems can emerge. In contrast, systems that are rigid and fixed cannot adapt to staying with the trouble, nor are they capable of confronting and taking responsibility for the horror and uncertainty of it all.
In fact, Dempster (1998) observed that Systems Theories from a rational-comprehensive approach (i.e engineering) tend to “bound that which has no boundaries” and their rigidity prevents them from responding to complex and dynamic social and environmental issues. Dempster’s work built on Autopoiesis and Cognition (1980), a cybernetic work in systems theory and philosophy of biology co-authored by Chilean biologists and philosophers Humberto Maturana and Francisco Varela. They make a case for the term autopoiesis, using the Greek prefix αυτό (auto: self) to describe the property identified in cells and organisms that allows them to maintain and renew themselves by self-regulating their composition and boundaries. Characteristics of autopoietic systems are:
Structure: autonomous units with self-defined boundaries
Terms of relation: closed (relations between parts)
Reproduction: Autopoietic: copy or self-production
Evolution: between systems
Goal: growth/development orientated
Balance: homeostasis
Trajectories: finite temporal trajectories
Pattern of organisation: closed (relations between parts)Dempster (1998)
Instead, Dempster proposes we recognise and plan also for sympoietic system (collectively-producing) and argues that they may be more capable of responding to sustainability challenges. It is important to note that Philosopher and anthropologist Anna Lowenhaupt Tsing (2017) clarifies that autopoiesis and sympoiesis are in generative friction rather than opposition to each other (p. M26) and that Margulis would have used ‘sympoietic’ at times to describe “interlocked and multileveled systemic processes of non-reductionist organization and maintenance” (p. M27). Contrary to autopoietic systems, sympoietic systems allow for flexible and adaptive planning that is capable of recognising “uncertain futures, synergistic possibilities, differing perspectives, and multiple values” (p.v). Characteristics of sympoietic systems are:
Structure: unbounded, complex, amorphous entities, lack self-defined spatial and temporal boundaries
Terms of Relation: cooperative
Reproduction: amorphously and collectively
Evolution: occurs within the system
Goal: evolutionary orientated
Balance: dynamic tension
Trajectories: potentially infinite temporal trajectories
Pattern of organisation: ajar, open to external information and merging, unpredictable, creativeDempster (1998)
Complex living systems are the original template from which the concept of sympoiesis was developed. Therefore ecosystems are an example of the ‘ideal’ sympoietic system. Other examples include: a forest, acommunity, an open group, common English, participatory processes and post-normal science (1998). Dempster does not use ‘sympoietic’ and ‘symbiotic’ interchangeably. Yet, the conditions for and characteristics of symbiosis overlap with several conditions for and characteristics of sympoietic systems. Both provide a counter-argument to the self or the individual as an autonomous unit and prove that novel capacities and capabilities emerge through the prolonged encounter of diversity, cooperation and complexity. This novel outcome or property is called an emergent property. It can either be a new species that demonstrates characteristics and functions that would not be present in the respective organisms had they not co-evolved, or the ability of small birds to confuse predators through their murmurations.
Emergence
Emergence is the outcome of a synthesis that displays novel characteristics and behaviours when compared to the individual parts of the system. Lichens, for example are an emergent property. Other examples, where you might encounter emergence are:
birds flocking
cells splitting
fungi forming
pheromone-dropping ants
mycelium communicating
fireflies synchronising
fish schooling
people organising into grassroots movements
the formation of galaxies and stars and planets
the evolution of life on earth
the folding of proteins
the assembly of cells
the crystallisation of atoms in a liquid
superconductivity of electrons in some metals
climate collapse
Potawatomi botanist Robin Kimmerer recounts in her book Braiding Sweetgrass (2013) the moment she first ‘tasted’ the missing language of her people when she stumbled upon the word Puhpowee in a treatise on the traditional uses of fungi by her people, written by Anishinaabe ethnobotanist Keewaydinoquay.
The makers of this word understood a world of being, full of unseen energies that animate everything. I’ve cherished it for many years, as a talisman, and longed for the people who gave a name to the life force of mushrooms. The language that holds Puhpowee is one that I wanted to speak. So when I learned that the word for rising, for emergence, belonged to the language of my ancestors, it became a signpost for me. (p. 44)
In European and Western geographies, emergence falls under two movements. On the one hand, Proto-emergentism (loosely joining the sciences, philosophy and theology) is based on the work of English philosopher and critic G.H. Lewes from 1875. Neo-emergence or Complexity Theory is rooted in cybernetics, solid state/ condensed matter physics, evolutionary biology, artificial intelligence and artificial life (De Wolf & Holvoet, 2005). Author, artist and social justice activist adrienne maree brown (2017) articulates the poetry-spirituality-physics-systems dynamics-evolutionary biology-of-it-all as:
The wonder and awe we experience being in the world is often the result of the recognition of how complex systems and patterns arise out of a multiplicity of relatively simple interactions. The world is full of beauty, magic, miracles, and patterns that induce wonder. Emergence is one of the best concepts I have learned for discussing this wow, this wonder (p.3).
brown (2018) echoes Dempster (1998) when she emphasises critical connections over critical mass, and building authentic relationships, listening with all the senses of the body and the mind (Brown, 2017, p.3). Such emergents display novel characteristics and behaviours compared to the the individual parts of the system (De Wolf & Holvoet, 2005, p. 3). Much like symbiogenesis, the continual intra-action of the individual parts over time is required.
Conclusion
I am interested in organisations, collectives and systems that require and facilitate the prolonged encounter of plurality and diversity, wherein friction becomes generative because cooperation is prioritised. I am curious about organisations, collectives and systems that respond to their sites with sensitivity and depth, changing, adapting or ending when it is time for other things to take their place. This is why the evolutionary process of symbiosis serves as the conceptual framework of this research to study how ‘the living together’ of critical environmental education and collective artistic practices may allow for the emergence of transformative ecopedagogies with/in/out of their institutional frameworks.
The purpose of this first reflection on Symbiosis was to introduce the reader to the idea of symbiosis as a biological and systemic process and to challenge certain hegemonic social imaginaries around competition, cooperation and survival. The second part of this essay (Publ. in Nov 2025) will continue an analysis on Emergence and how it may help us reconfigure artistic, curatorial, educational and institutional frameworks and practices.
If it is true that neither biology nor philosophy no longer support the notion of independent organisms in environments, that is, interacting units plus contexts/rules, then sympoiesis is the name of the game in spades. Bounded (or neoliberal) individualism amended by autopoiesis is not good enough figurally or scientifically; it misleads us down deadly paths.
Haraway (2016)
See also: Lichenizing Pedagogy
References
Brockman, J. (1996). The Third Culture: Beyond the Scientific Revolution. Simon & Schuster.
brown, a. m. (2017). Emergent Strategy: Shaping Change, Changing Worlds. AK Press.
Dawkins, R. (1976). The Selfish Gene. Oxford University Press
Dempster, B. (1995). System Stability and Implications for Sustainability [B. Sc Thesis].
Dempster, B. (1998). A Self-Organizing Systems Perspective on Planning for Sustainability [ M.E.S. Thesis].
Feldman, J. (Director).(2017). Symbiotic Earth: How Lynn Margulis rocked the boat and started a scientific revolution.
Garcés, M. (2012).Honesty with the real. Journal of Aesthetics & Culture, 4(1), 18820. DOI
Guerrero, R., Margulis, L., & Berlanga, M. (2013). Symbiogenesis: the holobiont as a unit of evolution. International Microbiology: The Official Journal of the Spanish Society for Microbiology, 16(3), 133–143. DOI
Haraway, D. (2016). Staying with the trouble: making kin in the Chthulucene. Duke University Press.
Hird, M. J. (2009). The Origins of Sociable Life: Evolution After Science Studies. Palgrave Macmillan London.
Huxley, J. (1942). Evolution: The Modern Synthesis., Allen & Unwin. London
Lowenhaupt Tsing, A. (2012). Unruly Edges: Mushrooms as Companion Species. Environmental Humanities, 1(1), 141–154. DOI
Lowenhaupt Tsing, A., Swanson, H., Gan, E., & Bubandt, N. (2017). Arts of living on a damaged planet. University Of Minnesota Press.
Machado de Oliveira Andreotti, V. (2021). Hospicing Modernity: Parting with Harmful Ways of Living. North Atlantic Books.
Margulis, L. (1998). Symbiotic planet: a new look at evolution. Basic Books.
Margulis, L., & Fester R. (Eds.). (1991). Symbiosis as a Source of Evolutionary Innovation: Speciation an Morphogenesis. MIT Press.
Margulis, L. (1995). Gaia is a tough bitch. In J. Brockman (Ed.), The Third Culture: Beyond the Scientific Revolution (p. 129–146). Simon & Schuster.
Maturana, H. R., & Varela, F. J. (1980). Autopoiesis and Cognition The Realization of the Living. Reidel.
Mbembe, A. (2003). Necropolitics. Duke University Press.
Meloni, M. (2014). How Biology Became Social, and What it Means for Social Theory. The Sociological Review, 62(3), 593–614. DOI
Palmer, L. A. (2023). The Lichen Museum. University of Minnesota Press.
Sapp, J. (1994). Evolution by Association: A history of Symbiosis. Oxford University Press.
Spencer, H. (1864). The Principles of Biology. Williams & Norgate.
